A eukaryotic cell cannot divide into two,
the two into four, etc. unless two processes alternate:
- doubling of its genome
(DNA) in S phase (synthesis phase) of the cell cycle
- halving of that genome
during mitosis
(M phase)
The period between M and S is called G1; that between S and
M is G2.
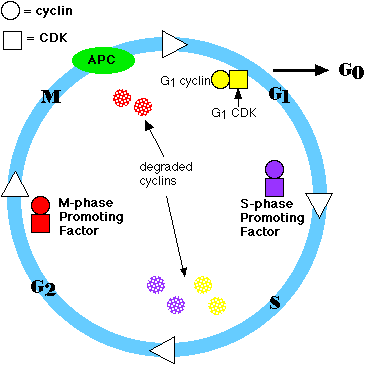 So, the
cell cycle consists of:
So, the
cell cycle consists of:
- G1 = growth and preparation of the chromosomes for
replication
- S = synthesis of DNA (and
centrioles) [see DNA Replication]
- G2 = preparation for
- M = mitosis
When a cell is in any phase of the cell cycle other than mitosis, it
is often said to be in interphase.
The
passage of a cell through the cell cycle is controlled by proteins in the
cytoplasm. Among the main players in animal cells are:
- Cyclins There are 3 groups:
- G1 cyclins
- S-phase cyclins
- M-phase cyclins
Their levels in the cell rise and fall
with the stages of the cell cycle.
- Cyclin-dependent kinases
(CDKs) Again, there are 3 groups:
- G1 CDKs
- S-phase CDKs
- M-phase CDKs
Their levels in the cell remain fairly
stable, but each must bind the appropriate cyclin (whose levels fluctuate) in
order to be activated. They add phosphate groups to a variety of protein
substrates that control processes in the cell cycle.
- The anaphase-promoting complex (APC) and other proteolytic enzymes.
The APC
- triggers the events leading to destruction of the cohesins
and thus allowing the sister chromatids to separate.
- degrades the mitotic (M-phase) cyclins
Steps in the cycle
- a rising level of G1 cyclins signals the cell to prepare
the chromosomes for replication
- a rising level of S-phase promoting factor (SPF) prepares
the cell to enter S phase and duplicate its DNA (and its centrioles)
- as DNA replication continues, one of the cyclins shared by G1
and S-phase CDKs (cyclin E) is destroyed and the level of mitotic cyclins
begins to rise (in G2)
- M-phase promoting factor (the complex of mitotic cyclins with
M-phase CDK) initiates
- assembly of the mitotic spindle
- breakdown of the nuclear envelope
- condensation of the chromosomes
- these events take the cell to metaphase of mitosis
- at this point, the M-phase promoting factor activates the anaphase
promoting complex (APC) which
- allows the sister chromatids at the metaphase plate to separate and move
to the poles (= anaphase), completing mitosis
- destroys the M-phase cyclins. It does this by conjugating them with the
protein ubiquitin which targets them for destruction by
proteasomes.
- turns on synthesis of G1 cyclins for the next turn of the
cycle
- degrades geminin, a protein that has kept the
freshly-synthesized DNA in S phase from being re-replicated before mitosis.
Meiosis and the Cell Cycle
The special behavior of
the chromosomes in meiosis I requires some special controls. Nonetheless, passage through the cell
cycle in meiosis I (as well as meiosis II, which is essentially a mitotic
division) uses many of the same players, e.g., MPF and APC. (In
fact, MPF is also called maturation-promoting factor for its role
in meiosis I and II of developing oocytes.
The cell has several systems for interrupting the cell cycle if
something goes wrong.
- A check on completion of S phase. The cell seems to
monitor the presence of the Okazaki fragments on the lagging strand during DNA
replication. The cell is not permitted to proceed in the cell cycle until
these have disappeared.
- DNA damage checkpoints. These sense DNA damage
- before the cell enters S phase (a G1 checkpoint);
- during S phase, and
- after DNA replication (a G2 checkpoint).
- spindle checkpoints. Some of these that have been discovered
- detect any failure of spindle fibers to attach to kinetochores
and arrest the cell in metaphase (M checkpoint - example)
- detect improper alignment of the spindle itself and
block cytokinesis
- trigger apoptosis
if the damage is irreparable.
All the checkpoints examined require the services of a
complex of proteins. Mutations in the genes encoding some of these have been
associated with cancer; that is, they are oncogenes.
This should not be surprising since checkpoint failures allow the cell to
continue dividing despite damage to its integrity.
p53
The p53
protein senses DNA damage and can halt progression of the cell cycle in both
G1 and G2. Both copies of the p53 gene must be mutated for
this to fail so mutations in p53 are recessive, and p53 qualifies
as a tumor suppressor gene.
The p53 protein is also a key player in apoptosis,
forcing "bad" cells to commit suicide. So if the cell has only mutant versions
of the protein, it can live on - perhaps developing into a cancer. More than
half of all human cancers do, in fact, harbor p53 mutations and have no
functioning p53 protein.
| A genetically-engineered adenovirus, called ONYX-015,
can only replicate in human cells lacking p53. Thus it infects,
replicates, and ultimately kills many types of cancer cells in vitro.
Clinical trials are now proceeding to see if injections of ONYX-015 can
shrink a variety of types of cancers in human patients.
|
In some way, p53 seems to evaluate the extent of damage to DNA, at least for
damage by radiation.
- At low levels of radiation, producing damage that can be repaired, p53
triggers arrest of the cell cycle until the damage is repaired.
- At high levels of radiation, producing hopelessly damaged DNA, p53
triggers apoptosis.
ATM
ATM (="ataxia telangiectasia mutated") gets its name from a human disease of that
name,
whose patients - among other things - are at increased risk of cancer. The ATM
protein is involved in
- detecting DNA damage, especially double-strand breaks;
- interrupting (with the aid of p53) the cell cycle when damage is found;
- maintaining normal telomere
length.
MAD
MAD (="mitotic arrest deficient") encodes a
protein that binds to each kinetochore until a spindle fiber (one
microtubule will do) attaches to it. If there is any failure to attach, MAD
remains and blocks entry into anaphase.
Mutations in MAD produce a
defective protein and failure of the checkpoint. The cell finishes mitosis but
produces daughter cells with too many or too few chromosomes (aneuploidy).
Aneuploidy
is one of the hallmarks of cancer cells suggesting that failure of the spindle checkpoint is a major step
in the conversion of a normal cell into a cancerous one.
Infection with the human T cell leukemia virus-1 (HTLV-1) leads
to a cancer (ATL = "adult T cell leukemia") in about 5% of its victims.
HTLV-1 encodes a protein, called Tax, that binds to the MAD protein
causing failure of the spindle checkpoint. The leukemic cells in these patients
show many chromosome abnormalities including aneuploidy.
A kinesin that moves the
kinetochore to the end of the spindle fiber also seems to be involved in the
spindle checkpoint.
Many times a cell will leave the cell cycle, temporarily or permanently. It
exits the cycle at G1 and enters a stage designated G0 (G
zero). A G0 cell is often called "quiescent", but that is probably
more a reflection of the interests of the scientists studying the cell cycle
than the cell itself. Many G0 cells are anything but quiescent. They
are busy carrying out their functions in the organism. e.g., secretion,
conducting nerve impulses, attacking pathogens.
Often G0 cells are terminally differentiated: they will never
reenter the cell cycle but instead will carry out their function in the organism
until they die.
For other cells, G0 can be followed by reentry into the cell
cycle. Most of the lymphocytes in human blood are in G0. However,
with proper stimulation, such as encountering the appropriate antigen, they can
be stimulated to reenter the cell cycle (at G1) and proceed on to new
rounds of alternating S phases and mitosis.
G0 represents not simply the absence of signals for mitosis but an
active repression of the genes needed for mitosis. Cancer cells cannot enter
G0 and are
destined to repeat the cell cycle indefinitely.
4 March 2003